Das Leben
Die Fotosynthese
Die wichtigste Reaktion der Welt
Die Fotosynthese umfasst die chemischen
Reaktionen, die mit Hilfe der Energie aus dem Sonnenlicht Wasser und
Kohlendioxid in Zucker und Sauerstoff umwandelt. Sie stellt nicht
nur alleine die Energieversorgung des höheren Lebens auf der Erde
sicherstellt (ohne die Fotosynthese wäre das Leben auf einfache
Einzeller, die von geochemischer Energie leben, beschränkt
geblieben, >> mehr),
sondern hat auch die fossilen Brennstoffe entstehen lassen, deren
Nutzung der Beginn der modernen Industriegesellschaft waren
(>>
mehr). Sie war es auch, die die Erde zu dem blau-grünen
Planeten gemacht hat, den wir heute kennen (>>
mehr).
Chloroplasten im Blatt
eines Laubmooses: Die Fotosynthese finden in
Chloroplasten
genannten Organellen in Algen und Pflanzen statt, die auf
eingewanderte Cyanobakterien zurückgehen (>>
mehr). Foto: Kristian
Peters, aus wikipedia, Artikel “Photosynthese” (abgerufen
27.1.2011).
Lizenz: >>
GNU FDL 1.2.
Um die Fotosynthese (50)
zu verstehen, wollen wir uns im Folgenden kurz einen Überblick über
ihren Ablauf verschaffen; und dann die einzelnen Schritte Punkt für
Punkt betrachten. Die Fotosynthese besteht aus einer Reihe von
>> Redoxreaktionen,
der dabei entstehende Elektronenfluss liefert Energie, mit dem
Organismen ihre universelle Energiewährung >>
ATP herzustellen und ein positiv geladenes Molekül namens NADP
zu reduzieren. Das dabei entstehende NAPDH wird genutzt, um aus
anorganischem Kohlendioxid den organischen, energiereichen
Zucker Glukose herzustellen, der der Ausgangspunkt für
alle weiteren biochemischen Reaktionen in der Zelle ist. Die
Elektronen für diesen Vorgang stammen aus Wasser, das bei der
Photosynthese oxidiert wird, als Abfallprodukt wird hierbei
Sauerstoff freigesetzt. Die Fotosynthese liefert also sowohl Energie
(in Form von ATP) als auch Glukose, das zentrale organische Molekül
des Stoffwechsels; und sie produziert Sauerstoff. Soweit zum
Überblick, kommen wir nun zu den Details:
Was ist eine Redoxreaktion?
Redoxreaktion steht für Reduktions-Oxidations-Reaktion: Das Wort
Oxidation bezeichnet historisch die >> Reaktion
einer Substanz mit Sauerstoff; Reduktion das Gegenteil, den
Entzug von Sauerstoff. Heute werden beide Begriffe allgemeiner
verwendet: von einer der Reduktion spricht man,
wenn ein Reaktionspartner ein oder mehrere Elektronen aufnimmt
(diesen Zusammenhang kann man sich so merken: da jedes Elektron eine
negative Ladung trägt (-1 geschrieben, mehr >> hier),
reduziert sich dadurch die Ladung des Moleküls, das ein Elektron
aufnimmt). Da bei chemischen Reaktionen die aufgenommenen Elektronen
irgendwo herkommen müssen, ist eine Reduktion immer mit einer Oxidation
verbunden, der Abgabe von Elektronen (Sauerstoff nimmt sehr leicht
Elektronen auf und führt daher zu einer Oxidation, was die
historische Bezeichnung erklärt. Sauerstoff ist daher ein starkes
Oxidationsmittel).
Eine Redoxreaktion besteht also immer aus zwei Teilreaktionen, eine
Reduktion ist mit einer Oxidation gekoppelt:
Reduktion: Molekül A (neutral) + zwei Elektronen
(zweimal -1) -> Molekül A (Ladung -2) (51)
Oxidation: Molekül B (neutral) -> Molekül B
(Ladung +2) + zwei Elektronen (Ladung zweimal -1)
Da sich das negativ geladene Molekül A und das positiv geladene
Molekül B anziehen, werden sie zu dem neuen Molekül AB reagieren:
Redoxreaktion: Molekül A + Molekül B -> Molekül
AB
Was man in der Formel nicht sieht: die Elektronen sind von Molekül
B zu Molekül A geflossen (werden aber nicht frei, weshalb sie in der
Formel nicht mehr auftauchen). Die Anziehungskraft eines Moleküls
für Elektronen kann in Form seines „Redoxpotenzials“
gemessen werden: Oxidationsmittel wie Sauerstoff, also Substanzen
mit starker Anziehungskraft auf Elektronen, haben ein positives
Redoxpotenzial; je höher das Redoxpotenzial ist, desto stärker ist
auch die Anziehungskraft auf Elektronen. Der bei Redoxreaktion
ausgelöste Fluss der Elektronen liefert – ganz ähnlich wie beim
Fluss >>
elektrischen Stroms – Energie, mit der andere chemische
Reaktionen angetrieben werden können.
Die Redoxreaktionen der Fotosynthese
Die Besonderheit bei der Fotosynthese sind zwei “Photosysteme”,
die unter dem Einfluss von Licht ihr Redoxpotenzial ändern
können. Außerdem können alle Organismen –
Cyanobakterien, Algen, Pflanzen -, die bei der Fotosynthese
Sauerstoff erzeugen (50),
etwas, was wir Menschen technisch auch gerne könnten: sie können
nämlich mit Hilfe von Sonnenlicht Wasser oxidieren und Wasserstoff
erzeugen. Dies geschieht am Photosystem II
(das aus historischen Gründen so heißt, denn es wurde als zweites
entdeckt – tatsächlich beginnt die Fotosynthese hier). Dieses ist
ein starkes Oxidationsmittel – so stark, dass es
das chemisch sehr stabile Wasser aufbrechen kann (dabei hilft ein
zum Photosystem II gehörendes Enzym, der “sauerstoffproduzierende
Komplex”) und Elektronen aus dem dabei entstehenden Wasserstoff
freisetzen. Diesen werden an einen “Elektronenakzeptor” am
Photosystem II gebunden. Fällt Licht auf diesen Elektronenakzeptor,
ändert sich dessen Redoxpotenzial (siehe Abbildung): das Photosystem
II verliert an Anziehungskraft für Elektronen, so dass diese wieder
freigesetzt werden (anders ausgedrückt – das Licht liefert die
Energie, die für die Freisetzung der Elektronen benötigt wird).
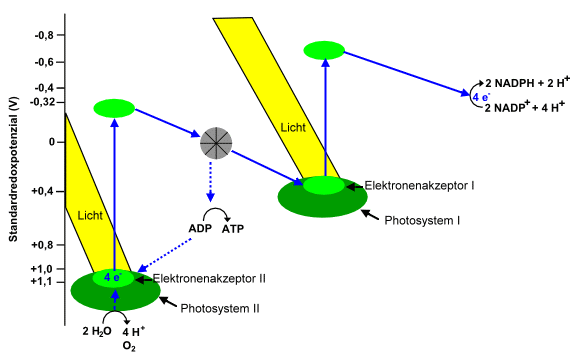
Die Lichtreaktion (Schritte 1 bis
4) der Fotosynthese (vereinfacht): Der Elektronenakzeptor
des Photosystems II ist als starkes Oxidationsmittel mit
Unterstützung des “sauerstoffproduzierenden Komplexes” in der Lage,
Elektronen aus Wasser aufzunehmen. Durch Licht angeregt, werden die
Elektronen wieder abgegeben und werden über eine Reihe chemischer
Reaktionen zum Photosystem I transportiert, wo sie durch
Lichteinfall wiederum angeregt werden und über eine Reihe weiterer
Reaktionen schließlich auf NADP+ übergehen, wobei NADPH entsteht,
dass die Dunkelreaktion, bei der Zucker entsteht, antreibt. Wegen
des Zickzackförmigen Verlaufs des Redoxpotenzials wird diese Form
der Darstellung, die auf den britischen Biochemiker Robin Hill
zurückgeht, auch “Z-Schema” genannt. Eigene Abbildung.
Über eine Reihe weiterer Redoxreaktionen (hier nicht im Einzelnen
dargestellt) fließen die vom Photosystem II freigesetzten Elektronen
zum Photosystem I (gelegentlich – siehe die gestrichelte blaue Linie
– auch zurück zum Photosystem II; bei diesem Weg wird ATP erzeugt).
Wichtig auf dem Weg zum Photosystem I ist vor allem, dass die
Energie aus dem Elektronenfluss dazu genutzt wird, Protonen (also
positiv geladene Wasserstoffmoleküle) durch eine Membran zu pumpen
– der graue Kreis soll diese “Pumpe” darstellen. Auch das
Photosystem I ändert sein Redoxpotenzial unter Lichteinwirkung, es
wird dann ein starkes Reduktionsmittel. Es
reduziert (wieder über eine Reihe hier nicht dargestellter
Reaktionen) schließlich positiv geladene Moleküle namens NADP
(Ladung +1) zu NADPH. Da alle diese Reaktionen von
der Änderung des Redoxpotenzials der Photosysteme durch Licht
abhängig sind, nennt man sie auch die Lichtreaktion.
Deren chemische Formel lautet also in der Summe:
2 H2O + 2 NADP+ -> 2 H+
+ 4e- + O2 + 2 NADPH
Dieses NADPH ist selber ein (wenn auch weniger starkes, siehe
Abbildung oben) Reduktionsmittel und reduziert Kohlendioxid zu dem
organischen Kohlenhydraten. Diese Reaktion kann auch ohne Licht
stattfinden, sie wird daher Dunkelreaktion
genannt. Ihr Ablauf wurde von den amerikanischen Biochemikern Melvin
Calvin und Andrew Benson aufgeklärt und ist daher als Calvin-Benson-Zyklus
bekannt (Melvin Calvin erhielt hierfür 1961 den Nobelpreis für
Chemie). Ihre Summenformel lautet:
CO2 + 2 NADPH + 2 H+ + 4e-
-> CH20 + H20 + 2 NADP+
Bei der Dunkelreaktion wird nicht nur das NADP+ wieder
regeneriert, sondern auch ATP in ADP umgewandelt, so dass auch
dieses "recycelt" wird. Die Licht- und die Dunkelreaktion ergeben
zusammen die Summenformel der Fotosynthese:
CO2 + H20 -> CH2O + O2;
oder besser, da letztlich der Zucker Glukose entsteht:
6 CO2 + 6 H20 -> C6H12O6
(Glukose) + 6 O2
Das Kohlendioxid für diese Reaktion stammt aus der Luft, und wird
über Spaltöffnungen in den Blättern aufgenommen, das Wasser wird von
Landpflanzen über die Wurzeln aus dem Boden aufgenommen. Was aus dem
Sauerstoff wird, steht >>
hier.
Ist
mehr Kohlendioxid gut für Pflanzen?
Kohlendioxid ist, wie oben gesehen, eine wichtige Zutat für die
Fotosynthese; andererseits aber auch ein Treibhausgas (>> mehr),
das zur Erderwärmung beiträgt (>> mehr).
Skeptiker, die die Warnungen von dem Klimawandel für übertrieben
halten, weisen gerne darauf hin, dass mehr Kohlendioxid in der Luft
Pflanzen besser wachsen lässt, und die Landwirtschaft daher vom
Klimawandel sogar profitieren könnte, und nicht, wie etwa vom
Weltklimarat befürchet (>> mehr),
ihre Produktivität zurückgehen würde. Als Beleg führen sie an, dass
etwa in Treibhäusern gezielt die Kohlendioxid-Konzentration erhöht
wird, um die Produktion zu steigern. Irrt hier der Weltklimarat?
Nein, denn die Welt ist kein Treibhaus: Die Steigerung der
Produktion tritt nur dann ein, wenn alle anderen Faktoren, vor allem
Luftfeuchtigkeit und Temperaturen gleich bleiben. Dies ist aber
“draußen” nicht der Fall, eine Welt mit mehr Kohlendioxid wäre
wärmer und in vielen Regionen während der Wachstumzeit auch
trockener. Das wichtige Enzym Rubisco ist aber “temperatursensitiv”,
seine Leistung geht (oberhalb eines bestimmten Niveaus) bei
steigenden Temparaturen zurück; und wenn es trockener ist, schließen
die Pflanzen ihre Spaltöffnungen, um die Wasserverluste zu
reduzieren. Durch diese nehmen sie aber auch Kohlendioxid auf, so
dass trotzt steigender Konzentration an Kohlendioxid in der Luft
weniger in die Pflanze gelangt. Durch die verringerte Verdunstung
steigt auch die Temperatur des Blattes, was wiederum die Leistung
der Rubisco verringert. Welche Effekte überwiegen bei steigender
Konzentration an Kohlendioxid? Dies versuchen Forscher in
sogenannten “free-air carbon dioxide enrichment”-Experimenten
(FACE – zu deutsch: Experimente mit Kohlendioxidanreicherung im
Freien) zu untersuchen. Diese Experimente, und ihre Verknüpfung mit
den zu erwartenden Klimabedingungen, sind allerdings noch nicht in
wichtigen Regionen wie den Tropen durchgeführt worden, und aufgrund
der geringen Zahl sind die Ergebnisse insgesamt mit Vorsicht zu
genießen – aber bisher deuten alle darauf hin, dass die
Produktivität trotzt steigender Kohlendioxid-Konzentration in einer
Welt des Klimawandels zurückgehen würde. So einfach, wie die
Skeptiker es sich vorstellen, sind die Zusammenhänge jedenfalls
nicht.
Dabei sind übrigens auch die mit Trockenheit besser
zurechtkommenden C4-Pflanzen (siehe folgenden Abschnitt) keine
Hilfe: Sie sind im Durchschnitt für die Ernährung von Mensch und
Tier schlechter geeignet, und unter den wichtigen Nahrungspflanzen
gehören nur Mais, Sorghum und Zuckerrohr zu den C4-Pflanzen.
Von der C3- und der C4-Fotosynthese
Im Detail ist der Calvin-Benson-Zyklus ein
komplexer Vorgang: Das Kohlendioxid wird durch eine Enzym namens
Ribulose-1,5-bisphosphat-Carboxylase (ein Name, der selbst
Wissenschaftlern zu lang ist und daher gerne zu Rubisco abgekürzt
wird) an einen Kohlenstoffakzeptor mit 5 Kohlenstoffatomen namens
Ribulose-1,5-bisphosphat gebunden, das Zwischenprodukt zerfällt dann
sehr schnell zu 2 Molekülen mit 3 Kohlenstoffatomen namens
3-Phosphoglycerat. Da diese erste fassbare Zwischenstufe 3
Kohlenstoffatome besitzt, spricht man bei dieser Grundform der
Fotosynthese auch von der “C3-Fotosynthese”. Aus zwei Molekülen
3-Phosphoglycerat wird der Zucker Glukose gebildet, aus zehn
Molekülen 3-Phosphoglycerat wird Ribulose-1,5-bisphoshat
regeneriert.
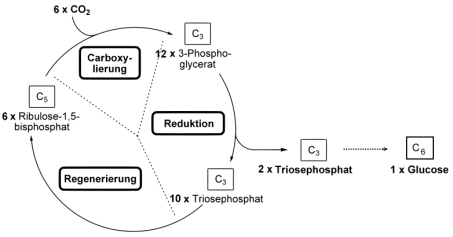
Der Calvin-Benson-Zyklus
(der Weg vom Triosephosphat zur Glukose gehört
nicht mehr hierzu, wurde aber zur Verdeutlichung hier eingefügt.
Neben
der Bildung von Glukose sind die Triosephosphate auch Ausgangsstoff
für Fette, Fett- und Aminosäuren). Abbildung verändert nach >>
wikipedia,
Stichwort
Calvin-Zyklus (abgerufen 25.7.2011). Gemeinfrei.
Das Enzym Rubisco hat jedoch eine Schwäche: Es kann nicht gut
zwischen Sauerstoff und Kohlendioxid unterscheiden. Gelegentlich
bindet es statt Kohlendioxid daher Sauerstoff an das
Ribulose-1,5-bisphosphat, und daher sinkt bei steigendem
Sauerstoffgehalt in der Luft die Fotosynthese. Dieser Vorgang heißt
Fotorespiration. Rubisco ist vermutlich bereits in der
sauerstofffreien frühen Zeit der Geschichte des Lebens entstanden,
damals spielte diese Schwäche keine Rolle; besonders starke
Auswirkungen muss die Fotorespiration in Zeiten hohen
Sauerstoffgehalts wie etwa im Karbon (>> hier)
gehabt haben. So haben die Pflanzen denn auch einen Ausweg erfunden,
der eigentlich ein Umweg ist – die “C4-Fotosynthese”. Bei dieser
wird das Kohlendioxid durch das Enzym PEP-Carboxylase an ein Molekül
mit 3 Kohlenstoffatomen, das Phosphoenolpyruvat (PEP) gebunden; über
eine Zwischenstufe entsteht als erstes stabiles Zwischenprodukt
Malat (ein Molekül mit vier Kohlenstoffatomen, daher
C4-Fotosynthese). PEP-Carboxylase kann im Unterschied zu Rubisco
zwischen Sauerstoff und Kohlendioxid unterscheiden; das entstehende
Malat “füttert” dann Rubisco mit Kohlendioxid, und dann setzt der
Calvin-Benson-Zyklus ein. Dieses Umweg kostet Energie (die
Herstellung von PEP verbraucht ATP), lohnt sich aber bei hohen
Sauerstoff- und niedrigen Kohlendioxidkonzentrationen dennoch, da
dadurch die Fotorespiration vermieden und so die Fotosyntheserate
erhöht.
C4-Pflanzen haben auch Vorteile bei trockenem Klima, da Pflanzen
bei Trockenheit ihre Spaltöffnungen zum Schutz gegen Wasserverluste
schließen müssen, und dadurch weniger Kohlendioxid aufnehmen können.
>>
Zur Biomasseproduktion durch die Fotosynthese siehe hier und
unten
Die Erzeugung von ATP
Wir hatten oben gesehen, dass die Energie der Redoxreaktionen
zwischen dem Photosystem II und I dazu genutzt wird, Protonen durch
eine Membran zu pumpen. Dadurch entstehen an der Membran eine
Spannungsdifferenz und ein Konzentrationsunterschied. Um diesen
Unterschied auszugleichen, sind die Protonen “bestrebt” (eine Folge
des zweiten Hauptsatzes der Thermodynamik – Ladungs- und
Konzentrationsunterschiede stellen “Ordnung” dar, und geordnete
Systeme streben zur “Unordnung”), wieder zurück auf die andere Seite
zu gelangen. Dies können sie mit Hilfe des Enzyms ATP-Synthase,
das die Energie aus dem Fluss der Protonen analog einer Wassermühle
zur Herstellung von ATP nutzt. In Chloroplasten ergeben 14 Protonen
drei ATP. (Mit diesem Mechanismus wird auch ATP aus dem Abbau von
Nahrungsstoffen erzeugt. Für die Aufklärung dieser “chemiosmotischen
Koppelung” erhielt der britische Biochemiker Peter
Mitchell im Jahr 1978 den Nobelpreis für Chemie.)
Thema Energie II –
Fotosynthese und globale Energieflüsse
Welcher Anteil der Sonnenenergie wird von der Fotosynthese in
Pflanzenmasse umgewandelt? Der Antwort auf diese Frage kann man
sich auf zwei Wegen annähern: Zum einen kann man die
Energieumwandlung während der Fotosynthese betrachten, zum anderen
versuchen, die globale Produktion von Pflanzenmasse zu messen.
Fangen wir mit der ersten Möglichkeit an, der Energieumwandlung
während der Fotosynthese: Die Energie für die Lichtreaktion stammt
aus der Sonnenstrahlung; Sonnenstrahlung besteht aus Lichtquanten
(>> mehr).
Deren Energiegehalt hängt von der Wellenlänge des Lichts ab; die
fotosynthetisch aktive Strahlung (also die, die die Lichtreaktionen
antreibt) hat eine Wellenlänge von 400 bis 700 Nanometern,
Lichtquanten eines Durchschnittswerts von 550 Nm einen Energiegehalt
von 3,61 x 10-19 J (52).
Für die Fixierung eines mols Kohlendioxid werden nach experimentellen
Befunden 9 – 10 mol Photonen gebraucht, und diese haben einen
Energiegehalt von 2.065 kJ. Der Energiegewinn des in Zucker
gebundenen Kohlenstoffatoms im Vergleich zum Kohlendioxid beträgt
465 kJ, damit beträgt die Wirkungsgrad dieses Prozesses 465/2.065 =
0,225 oder 22,5 Prozent. Dieser Wert gilt aber nur die
fotosynthetisch aktive Strahlung der oben genannten Wellenlänge,
die nur etwa 43 Prozent der gesamten Sonnenstrahlung ausmacht; vom
Sonnenlicht insgesamt können daher maximal (22,5 x 0,43) etwa 10
Prozent genutzt werden. In der Praxis werden auch diese Werte nicht
erreicht: Ein Teil der Photonen wird von den Blättern reflektiert,
andere treffen nicht auf ein Photosystem, ein Teil der angeregten
Elektronen kann von den Enzymen nicht sofort verarbeitet werden und
die Energie geht als Abwärme verloren, ein Teil der Energie wird
durch die Fotorespiration verbraucht. Im besten Fall erreichen
Pflanzen kurzfristig bei optimalen Bedingungen daher einen
Wirkungsgrad von der Umwandlung von Sonnenlicht in gespeicherte
chemische Energie (Nettofotosynthese) von 6 bis 8 Prozent.
Optimale Bedingungen sind aber selten, auf großen Teilen der
Erdoberfläche sind Wassermangel oder zu niedrige/zu hohe
Temperaturen limitierende Faktoren, anderswo, etwa im
Amazonas-Regenwald, die Mineralienzufuhr. Daher liegt der im
Durchschnitt von der Fotosynthese in chemische Energie umgewandelte
Anteil der Sonnenenergie eher bei 1,5 Prozent.
Bestätigt wird dieser Wert vom anderen Ansatz, die Messung der
weltweiten Produktion von Biomasse. Diese wird auf verschiedene
Weisen in drei gebräuchlichen Einheiten ermittelt – Trockenmasse
(Biomasse ohne das darin enthaltene Wasser), enthaltener
Kohlenstoff oder Energiegehalt (mit folgenden Umrechnungsfaktoren:
2,2 Tonnen Trockenmasse entsprechen etwa 1 Tonne Kohlenstoff und
38,5 GJ Energie).
Gemessen werden kann die Bruttoprimärproduktion
– die gesamte mittels Fotosynthese in einem Jahr produzierte
Pflanzenmasse (entsprechend der oben genannten Nettofotosynthese),
die Nettoprimärproduktion – hier wird von der
Bruttoprimärproduktion der Eigenverbrauch der Pflanzen abgezogen
(die Zellatmung, die nicht mit der Fotorespiration während der
Fotosynthese verwechselt werden darf – die Zellatmung treibt die
Lebensvorgänge der Pflanze an, >> mehr),
oder die Nettoproduktion von Ökosystemen, bei der
von der Nettoprimärproduktion noch der Verbrauch durch die in einem
Ökosystem lebenden Mikroben und Tiere abgezogen wird.
Am gebräuchlichsten ist die Nettoprimärproduktion (NPP). Solche
globalen Werte können allerdings selbst mit modernen
Satellitentechniken (53)
nicht sehr genau gemessen werden; genaue Messungen am Boden zeigen
regelmäßig höhere Werte als die Satellitenmessungen. Hier liegt ein
Unsicherheitsfaktor, die jährliche Nettoprimärproduktion könnte also
höher liegen als die durchschnittlich 55 Gigatonnen
Kohlenstoff (121 Gigatonnen Trockenmasse) an Land
und 48,5 Gigatonnen Kohlenstoff (107
Gigatonnen Trockenmasse) im Ozean, die sich aus
Satellitenmessungen ergeben. Die Bruttoprimärproduktion oder
Nettofotosynthese beträgt global etwa das doppelte der
Nettoprimärproduktion, Algen und Pflanzen produzieren also
mindestens 456 Gigatonnen Trockenmasse im Jahr,
dem entspricht ein Energieinhalt von 8.000 EJ oder einer
durchschnittlichen Leistung der Fotosynthese von 250 TW (54)
– das ist ein Vielfaches dessen, was die Menschheit an technischer
Energie produziert (>> hier).
>>
Thema Energie III – Energieflüsse des Ökosystems Erde
Bionik I – Die Arbeit am
künstlichen Blatt
(Bionik)
Die Natur hat mit der Fotosynthese etwa erfunden, war
wir Menschen gerne könnten: Sie nutzt die Sonnenenergie, um damit
umweltfreundlich Energie herzustellen. Der Mechanismus ist
kompliziert, und unterscheidet sich damit von der Photovoltaik, dem
technischen Weg, aus Sonnenlicht Strom zu erzeugen, der eigentlich
ganz einfach ist (>>
hier) und – abgesehen von den Elektronen – ohne bewegte Teile
auskommt. Solarzellen haben zudem einen höheren Wirkungsgrad als
Pflanzen, sie können über 20 Prozent – und im Labor bereits 40
Prozent – der im Sonnenlicht enthaltenen Energie in Strom umwandeln.
Aber Solarzellen sind auch aufwändig in der Herstellung und
gefährlich als Abfall, Pflanzen dagegen wachsen von alleine und sind
essbar. Kein Wunder also, dass die Fotosynthese viele Forscher
anregt, die an neuen Formen der Energiegewinnung arbeiten.
Interessant ist dabei vor allem die Nutzung von einfachen
Farbstoffen statt teurer Siliziumkristalle, um aus Sonnenlicht Strom
zu erzeugen, und die Spaltung von Wasser in Wasserstoff und
Sauerstoff, gilt Wasserstoff doch als idealer Energiespeicher einer
Zukunft, die auf erneuerbare Energien setzt (>>
hier). Beide Forschungsrichtungen zeigen erste Erfolge: Für
seine Arbeiten an Solarzellen, die mit Farbstoffen arbeiten, erhielt
der Schweizer Chemiker Michael Grätzel im Jahr 2010 den
Millennium-Technologie-Preis (>>
hier, den Preis nannte die Zeitschrift GEO “eine Art
Nobelpreis für Ingenieure); an der Spaltung von Wasser nach dem
Vorbild der Fotosynthese arbeitet eine Arbeitsgruppe unter dem
Chemiker Daniel Nocera am Massachusetts Institute of Technology
(MIT), mehr: >> MIT
>> Technology
Review, (beide englischsprachig). Andere Forscher arbeiten
bereits daran, Noceras Katalysator mit Solarzellen oder besser noch
einfachen Farbstoffen zu einem “künstlichen Blatt” zu verbinden. Die
amerikanische Chemikerin Angela Belcher hofft sogar, diese
künstlichen Blätter von Viren herstellen lassen zu können (>>
mehr), die lichtfangende Farbstoffe und wasserspaltende
Katalysatoren auf engsten Raum zusammenbringen. Im Labor
funktioniert das schon, jedoch mit einem für die praktische
Anwendung viel zu teuren Katalysator. Auch die EU fördert Projekte
zur solaren Wasserstofferzeugung (>>
hier).