
Das Leben
Die Entfaltung des Lebens auf der Erde – 2
Der Weg zum vielzelligen Organismus
Die Eukaryoten entstehen
Eukaryoten kommt von griech. eu, “echt” und karyotos, “nussartig”, “Kern” – gemeint ist der Zellkern, der alle Eukaryoten von den Bakterien und Archaeen unterscheidet, die auch als Prokaryoten bezeichnet werden (Pro- von griech. pro, “vor”, also Organismen “vor dem Zellkern”). In diesem Zellkern der Eukaryoten, der durch eine Membran vom übrigen Zellinhalt abgegrenzt ist, befindet sich die DNS. Eukaryoten sind viel größer als Prokaryoten – im Durchschnitt 10.000 bis 15.000 Mal; in ihrem Zellkern befindet sich bis 1.000 Mal mehr DNS. Diese liegt auch nicht wie bei den Bakterien als einfacher Ring vor, sondern ist mit Proteinen zu Chromosomen verpackt (>> mehr). Daneben gibt es aber noch weitere Unterschiede: Typisch für Eukaryoten sind auch die Organellen, die ähnlich den Organen im Körper spezielle Aufgaben in der Zelle übernehmen. Dazu gehören etwa die Mitochondrien, die “Kraftwerke der Zellen”, in denen die universelle Energiewährung ATP hergestellt wird, oder die Chloroplasten der Pflanzen, in denen die Photosynthese stattfindet. Auch in ihrem Verhalten unterscheiden sie sich: Eukaryotenzellen können beispielsweise andere Zellen “fressen” – diese Fähigkeit heißt Phagocytose; diese Fähigkeit ist etwa in den Zellen unseres Immunsystem anzutreffen.
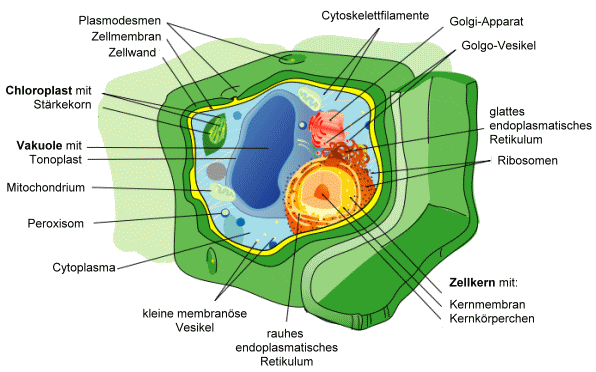
Eukaryoten- (hier: Pflanzen-)Zelle: Die DNS ist weitgehend im Zellkern eingeschlossen; die Zelle besitzt mehrere Organellen wie Mitochondrien und (bei Pflanzen) Plastiden wie die Chloroplasten. Siehe Text. Abbildung von Mariana Ruiz Villarreal, aus wikipedia commons, >> Abb. Pflanzenzelle (abgerufen 23.7.2008), Public Domain
{kind=link}
Prokaryoten und Eukaryoten unterscheiden sich also deutlich, und Zwischenstufen gibt es nicht. Wenn irgendwo in der Evolution von “missing links” (fehlenden Verbindungen) gesprochen werden kann, dann hier. Auch genetische Untersuchungen, etwa mit molekularen Uhren, haben zunächst fragwürdige Ergebnisse ergeben: Demnach wären die Eukaryoten 5 Milliarden Jahre alt – älter also als die Erde. Wenn wir nicht annehmen wollen, dass sie aus dem Weltall auf die Erde gekommen sind (was kaum jemand ernsthaft tut), kann die molekulare Uhr nicht stimmen – dies wäre damit zu erklären, dass es Zeiten in der Vergangenheit gab, in der die Evolution schneller verlaufen wäre. Genauere Untersuchungen der Gene haben dann auch noch ergeben, dass die Eukaryoten-DNS sowohl Gene enthält, die für Bakterien, als auch solche, die für Archaeen charakteristisch sind. Genetisch sind Eukaryoten Mischwesen; sie sind vermutlich nicht durch Darwinsche Evolution, sondern durch eine Art große Genfusion entstanden.
Dass so etwas grundsätzlich möglich ist, zeigt ein Blick auf den Bakterien-Sex: Bakterien vermehren sich durch Teilung, können aber auch mit Hilfe sogenannter “Sexpili” DNS auf andere Bakterien übertragen – und zwar über Artgrenzen hinweg (Biologen sprechen vom horizontalen Gentransfer – im Unterschied zum vertikalen Gentransfer, bei dem Gene von Eltern auf die Nachkommen übertragen werden). Diese Fähigkeit haben, wie die Ergebnisse genetischer Untersuchungen nahelegen, offenbar bereits die frühen Prokaryoten gehabt, und sogar in wesentlich größerem Umfang als heute. Vermutlich nahm die Bedeutung im Laufe der Zeit ab, als die Erbanlagen immer “perfekter” wurden – zufällige Genübertragung hat dann mit größerer Wahrscheinlichkeit negative Auswirkungen. Zur Zeit der Entstehung der Eukaryoten galt das offenbar noch nicht, damals muss es sogar zu Fusionen des kompletten Erbguts von Lebewesen gekommen sein. Dies zeigt die Entstehung der Organellen. Nach der Endosymbiontentheorie, die im Jahr 1967 von der amerikanischen Biologin Lynn Margulis vorgestellt (22) wurde, wären beispielsweise die Mitochondrien einst eigenständige Bakterien gewesen. Diese Bakterien könnten beispielsweise in der Lage gewesen sein, Sauerstoff zu nutzen, und hätte ihren Energieüberschuss im Austausch gegen andere Stoffe einem anderen Organismus zur Verfügung gestellt. Solche “symbiotischen” Beziehungen sind bei Prokaryoten nicht selten, sondern eher die Regel. Schließlich wäre in diesem Fall das Zusammenleben so eng geworden, dass schließlich eine Zelle in die andere aufgenommen wurde – dadurch wurde die Kontaktfläche zum Austausch größer.
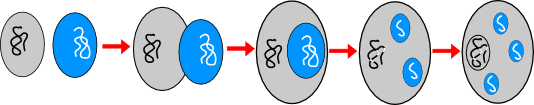
Die Entstehung der Eukaryoten nach der Endosymbiontentheorie: Ein Bakterium, dass Sauerstoff nutzen kann (blau), wird von einem anderen Bakterium aufgenommen, aber nicht verdaut, so dass es sich weiter vermehren kann. Im Laufe der Zeit verliert es einen Teil seines Erbguts und die Fähigkeit selbstständig zu überleben. Es wird zu einem Teil der späteren Eukaryotenzelle, dem Mitochondrium. (Anmerkung: Zur Zeit der Entwicklung der Endosymbiontentheorie war der Unterschied zwischen Archaeen und Bakterien noch nicht bekannt. Heute geht man davon aus, dass es ein Archaeon war, das ein Bakterium aufgenommen hat.) Zur Entstehung der Membran, die den Zellkern mit der DNS umgibt, gibt es verschiedene Hypothesen. Eine davon lautet, dass diese entstanden ist, da so der Transport von m-RNS zur den Ribosomen, an denen die Proteinbiosynthese stattfindet, kontrolliert werden kann. Siehe Erläuterungen im Text.
Diese Endosymbiose führte dazu, dass den Zellen wesentlich mehr Energie zur Verfügung stand als zuvor: Die Energiegewinnung bei Archaeen und Bakterien ist ja, wie wir gesehen haben, auf einen Protonengradienten über die Zellmembran angewiesen. Das Größenwachstum ist damit aus geometrischen Gründen beschränkt: die Oberfläche wächst bei einem kugelförmigen Prokaryoten im Quadrat des Radius, das Volumen aber im Kubik. Das heißt, je größer die Zelle wird, desto kleiner wird die Oberfläche im Verhältnis zum Volumen. Die Oberfläche ist aber die energieproduzierende Zellmembran; eine größer werdende Zelle wird also je Volumeneinheit schlechter mit Energie versorgt. Das dürfte der Grund sein, warum die Archaeen und Bakterien zwar eine sehr große biochemische Vielfalt entwickelt haben, aber im Laufe der gesamten Erdgeschichte immer klein geblieben sind. Mit der Endosymbiose und der Entwicklung der Mitochondrien war diese energetische Beschränkung aber aufgehoben, denn jetzt fand die Energiegewinnung an den Mitochondrienmembranen statt; und eine größer werdende Zelle könnte einfach ein größere Anzahl von Mitochondrien ausbilden, so dass die Energieproduktion pro Volumeneinheit gleich blieb oder sogar ansteigen konnte (23): die strukturelle Beschränkung des möglichen Zellwachstums wurde aufgehoben, die Zellen konnten größer werden.
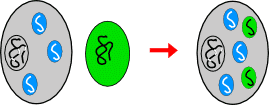
Die Entstehung der Chloroplasten
nach der Endosymbiontentheorie II:
Eine eukaryotische Zelle nimmt ein Cyanobakterium auf; aus
diesem wird
der Chloroplast einer einzelligen Alge, aus der die höheren Pflanzen
hervorgehen sollten. Eigene Abbildungen.
Eine ähnliche Symbiose sollte später zwischen Cyanobakterien, die zur Fotosynthese fähig waren, und einem Eukaryoten entstehen; aus den Cyanobakterien wurden so die Plastiden einer Alge. Diese Symbiosen brachten offenbar beiden Partnern so viel Nutzen, dass sie im Laufe der Zeit ihre Fähigkeiten, ohne den Partner zu überleben, verloren und miteinander “verschmolzen”. Aber noch heute besitzen Mitochondrien und Plastiden eigenes genetisches Material und teilen sich selbstständig; der Nachweis der Ähnlichkeit ihres Erbmaterials mit ihren mutmaßlichen Bakterienvorgängern (bei den Mitochondrien mit Alpha-Proteobakterien, bei Chloroplasten mit Cyanobakterien) verhalf der Endosymbiontentheorie zum Durchbruch – zumindest was Mitochondrien und Plastiden angeht. (Ähnlich entstanden später auch andere biologische Strukturen: Im Darm australischer Termiten lebt ein Einzeller, der diesen bei der Verdauung von Zellulose hilft. Seine Oberfläche ist mit “Haaren” bedeckt – die gar keine Haare, sondern fadenförmige Bakterien (Spirochäten), die koordiniert schlagen.) Die “Wirtszelle” war nach Ansicht der meisten Forscher ein Archaeon (wie die Archaeen im Singular heißen), was das genetische Mischwesen der heutigen Eukaryoten erklären würde. Wie aber könnte der namengebende Zellkern entstanden sein?
Lynn Margulis ging davon aus, dass auch der Zellkern die Folge eines Verschmelzens verschiedener Bakterien ist: Archaeen verschmolzen mit schwimmenden Bakterien ähnlich den heutigen Spirochaeten, die dort einige heute in Eukaryotenzellen zu findende Strukturen bildeten. Andere Wissenschaftler glauben, dass das Erbmaterial aus symbiotischen Partnerzellen beim Übergang in das der Wirtszelle derart negative Wirkungen hatte, dass im Verlauf der Evolution ein schützender Zellkern gebildet wurde. Dann stellt sich aber die Frage, warum die Bakterien sich nicht ebenfalls vor horizontalem Gentransfer schützen? Eine interessante alternative Erklärung schlugen Bill Martin und Eugene Koonin, von denen wir oben bereits eine Theorie zur Entstehung von Bakterien und Archaeen gesehen haben, vor (24): Eukaryoten besitzen in ihren Genen nichtcodierende DNS-Abschnitte, "echte" Introns. Bei Prokaryoten gibt es diese nicht, wohl aber die vermutlichen Vorgänger, "mobile" Introns (diese kommen nicht in Genen, sondern in anderen Bereichen der DNS vor, zerteilen daher keine Gene und sind damit keine "echten" Introns). Diese mobilen Introns sind einfach mobile, "egoistische" Stückchen von DNS: sie codieren Proteine wie die reverse Transkriptase, die aus RNS wieder DNS herstellt, die sich selbst wieder in die DNS der Wirtszelle einbaut. (Prokaryoten können offenbar eine gewisse Menge solcher DNS tolerieren, so dass es typischerweise etwa 30 mobile Introns in einem Bakteriengenom gibt.) Bei der Endosymbiose seien nun, so Martin und Koonin, vermutlich solche mobilen Introns aus den eingewanderten Bakterien in die Wirtszelle freigesetzt worden und hätten sich an der Entstehung des Eukaryoten-Genoms beteiligt. So wären die zerstückelten Eukaryoten-Genome erklärbar. Die von Introns sequenzierten Sequenzen müssen vor der Proteinbiosynthese entfernt werden, um keine Ressourcen an die Herstellung nicht funktionsfähiger Proteine zu verschwenden. Dieses “Herausschneiden” erfolgt durch "Spleißsomen" (die sich vermutlich aus den bakteriellen RNS-Scheren, mit denen sich die mobilen Introns aus der RNS herausschneiden, entwickelt haben); dieser Vorgang jedoch langsamer als die Proteinbiosynthese. Die Zellmembran wäre nach der Hypothese von Martin und Koonin der Weg, mit dem die Eukaryoten-Zelle den Transport der m-RNS mit den Introns von der DNS (im Zellkern) zu den Ribosomen außerhalb des Zellkerns (wo die Proteinbiosynthese stattfindet) zu kontrollieren und sicherzustellen, dass nur “reife”, gespleißte m-RNS ohne Introns zur Herstellung von Proteinen verwendet wird. Die Kernmembran ähnelt einer bakteriellen Membran; vermutlich wurden die Gene für die bakterielle Lipidherstellung für ihre Herstellung genutzt. Die übrigen Membranstrukturen wie die "Kernporen", die die Durchlässigkeit der Membran gewährleisten, werden sowohl von Bakterien- als auch Archaeengenen codiert, was dafür spricht, dass die Kernmembran sich erst nach der Endosymbiose ausgebildet hat.
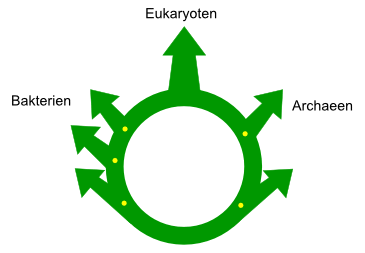
Der “Ring des Lebens”:
Aufgrund von horizontalem Gentransfer
und Genomfusionen lässt sich für die Entstehung der Eukaryoten
kein “Stammbaum” zeichnen, besser lässt sich die Entfaltung
des Lebens mit einem Ring darstellen: Bakterien und Archaeen
sind aus einem gemeinsamen Vorfahren entstanden, danach
sind die Genome bei der Entstehung der Eukaryoten teilweise
wieder vermischt worden. Eigene Abbildung nach >>
Nick Lane,
Life Ascending, figure 4.4.
Die Endosymbiontentheorie bedeutet, dass alle heutigen Lebewesen die beiden wichtigsten biochemischen Vorgänge – die Fotosynthese zum Aufbau organischer Substanz aus Sonnenlicht und die langsame Verbrennung von Sauerstoff zur Gewinnung von Energie aus dieser organischen Substanz – gewissermaßen in ihnen lebenden Bakterienkolonien verdanken. Unser Stoffwechsel beruht in wesentlichen Bestandteilen auf chemischen Verfahren, die bereits bei Bakterien und Archaeen anzutreffen sind. (Im Detail ist die Geschichte manchmal sogar noch komplizierter: Bei manchen Pflanzen besitzen die Chloroplasten eine doppelte Membran – sie gehen möglicherweise auf eine Symbiose mit eingewanderten Algen zurück, die ihrerseits durch die Aufnahme von zur Fotosynthese fähigen Cyanobakterien in eine Archaeenzelle entstanden sind, also auf eine doppelte Einwanderungsgeschichte.)
Wie der Sex auf die Welt kam
Prokaryoten-Sex besteht, wie wir oben gesehen haben, aus horizontalem Gentransfer; alle Eukaryoten treiben “echten” Sex: Dieser besteht darin, dass spezialisierte Fortpflanzungszellen, die Keimzellen – beim Menschen große, unbewegliche Eizellen und kleine, bewegliche Spermien –, miteinander verschmelzen und den Tochterorganismus bilden. Zur Bildung der Keimzellen wird das Erbgut der Elternzelle zunächst halbiert, so dass jede Keimzelle nur die Hälfte es elterlichen Erbguts enthält (25). Bei der Verschmelzung entsteht wieder ein komplettes Erbgut, das je zur Hälfte aus den beiden Keimzellen stammt. Da echter Sex bei allen Eukaryoten vorkommt (die wenigen Ausnahmen, die ohne Sex überleben können (26), haben diesen Weg später entwickelt), kann man vermuten, dass der Sex sich sehr schnell entwickelt hat und bereits existierte, als die großen Linien der Eukaryoten, die etwa zu Algen, Pflanzen, Pilzen und Tieren führen, sich getrennt haben. Dafür sprechen auch andere Gemeinsamkeiten aller Eukaryoten, die nicht auf Archaeen oder Bakterien zurückgehen, wie das dynamische innere Zellskelett, innere Membranen und der Zellkern. Die einfachste Erklärung hierfür ist Sex: die ersten Eukaryoten haben sich untereinander gekreuzt; alle Eukaryoten gingen also auf eine kleine, sich miteinander fortpflanzende "Urpopulation" zurück, die genau diese gemeinsamen Merkmale entwickelt hat.
Warum aber ist der Sex entstanden? Die Biologen haben diese Frage lange diskutiert, denn Sex hat einige auf den ersten Blick erkennbare Nachteile. Dazu gehört etwa der hohe Aufwand, der mitunter zur Partnersuche getrieben werden muss, oder dieser: Da zur Entstehung eines neuen Organismus zwei Keimzellen verschmelzen müssen, wird die Zahl der Tochterzellen im Vergleich zur einfachen Zellteilung halbiert. Diesem Nachteil muss ein noch größerer Vorteil gegenüberstehen, sonst hätte sich der Sex in der Evolution nicht durchgesetzt. Er hat vermutlich mit der DNS zu tun, die in Eukaryoten in Chromosomen organisiert ist. Bei der Entstehung der Keimzellen und der späteren Verschmelzung zum Tochterorganismus, werden Teilstücke der Chromosomen gegeneinander ausgetauscht. Biologen nennen diesen Vorgang “Rekombination”; dabei werden die Erbanlagen der jeweiligen Elternzellen miteinander gemischt. Diese Rekombination, so vermutete der Freiburger Zoologe August Weismann schon 1904, erhöhte die genetische Vielfalt, auf die die natürliche Selektion wirken kann. Damit würden “gute” Genkombinationen auf den Chromosomen langfristig zunehmen, “schlechte” aus dem Genbestand eliminiert. Später kamen von anderen Forschern weitere Überlegungen dazu: Die Durchmischung des Erbguts dürfte auch ein wirksamer Schutz vor Parasiten aller Art zu sein, die bei der ansonsten herrschenden genetischen Monokultur ein leichtes Spiel hätten. (Im reichen Westen vergessen wir dies ja gerne: Auf der Welt sterben viel mehr Menschen an Parasiten als etwa an Raubtieren.) Diese Erklärungen ist nach wie vor anerkannt, allerdings stellte sich ein theoretisches Problem: die langfristige Zunahme der “guten” Erbanlagen mag gut für die Art sein, aber die Evolution wirkt ja immer auf Individuen. Welchen Vorteil aber hat ein Individuum von der Rekombination? Auch hier liegt die Antwort bei den Chromosomen. Ohne Rekombination könnte die natürliche Auslese immer nur ganze Chromosomen betreffen: Gäbe es auf einem Chromosom eine Mutation, die ein Gen derart vorteilhaft machen würde, dass dieses Chromosom alle anderen Varianten verdrängen würde, ginge die genetische Vielfalt der Art zurück, da alle anderen Gene auf den anderen Varianten des Chromosoms ebenfalls verdrängt würden. Die Rekombination hilft also der Evolution, einzelne Gene zu "erkennen", indem sie starre Genkombinationen aufbricht. “Gute” Gene können durch Rekombination von einem “schlechten” Chromosom gerettet werden. Von diesem Mechanismus, so konnten Populationsgenetiker zeigen (27), profitieren auch die Individuen. Der Biochemiker Nick Lane (135) vermutet, dass die Rekombination besonders bei großen Genomen und hoher Mutationsrate überlegen ist. Durch die Introns aus den eingewanderten späteren Mitochondrien ist das Genom der Wirtszelle aber gewachsen; und ihr Einbau bedeutete eine höhere Mutationsrate – das Genom war genau den Faktoren ausgesetzt, bei denen die Rekombination besser war; die Endosymbiose könnte als auch der Auslöser dafür gewesen sein, dass sich der Sex entwickelt hat.
Die zweite Frage im Zusammenhang mit der Sexualität ist die, warum es zwei Geschlechter gibt. Die Vorteile der Sexualität wären auch mit einem Geschlecht zu haben (es müsste nur die Selbstbefruchtung verhindert werden), und die Anzahl möglicher Sexualpartner würde sich verdoppeln. Auch dies könnte mit der Endosymbiose, die zur Entstehung der Mitochondrien führte, zusammenhängen: In der Regel werden die Mitochondrien nämlich nur von einem der beiden Geschlechter vererbt (beim Menschen immer von der Mutter). Da die Mitochondrien einen großen Teil ihres eigenen Genoms verloren haben, was sie ja erst zu so guten Energielieferanten machte (23), muss ihr verbliebenes Genom mit dem der Wirtszelle kooperieren. Die hierfür notwendige Anpassung wird durch die Vererbung über nur ein einziges Geschlecht (uniparentale Vererbung) erleichtert: die Varianz (Vielfalt) zwischen den befruchteten Eizellen ist dabei deutlich größer, und gute Varianten sind für die Selektion besser "erkennbar". Diese Kombination – uniparentale Vererbung der Mitochondrien in einem Organismus, der aufgrund der Rekombination eine neue Zusammensetzung des Genoms hat, ist aber nur möglich, wenn es mindestens zwei Geschlechter gibt. Dafür sollten wir dankbar sein, denn wenn der Aufwand für die Partnersuche sicher ein Preis ist, der für den Sex zu zahlen ist, so machte er die Welt auch schöner: